
De cirkel in het midden stelt de versmelting voor van de mannelijke zaadcel met de vrouwelijke eicel. Deze nieuwe cel heeft weer het complete aantal (32) chromosomen en noemt men de kiemcel. Uit deze ene cel ontstaan twee cellen, uit de twee cellen ontstaan vier cellen, daarna acht, vervolgens zestien, enz. Dat gaat zo door tot dat cellen zich gaan specialiseren en dat gebeurt al heel snel.
Op de chromosomen liggen de genen, de dragers van de erfelijke eigenschappen. De erfelijkheid is niets anders dan via de geslachtscellen doorgeven van erfelijke eigenschappen. Van het aantal ingebrachte chromosomen komt dus de helft (16) van de vader en de helft (16) van de moeder, zodat de nieuwe cel weer 32 chromosomen telt
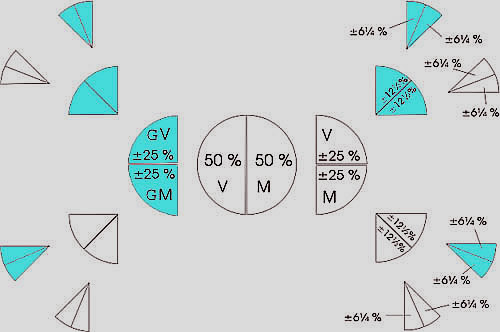
De blauwe segmentparen zijn telkens de grootouders van de dar uit het voorgaande segment. De dar heeft géén vader, maar alleen een moeder. De dar krijgt slechts via zijn moeder de genen door van zijn 2 grootouders. Het segment van de moeder van de dar kan daarom gevoeglijk worden weggelaten.
Elke koninginnenteler is er mee vertrouwd, dat koningin en dar gelijke bijdragen leveren aan de erfelijke basis van de bij. Maar niet iedereen zal zich realiseren dat de erfelijkheid van eigenschappen ver teruggrijpt en dat er zoveel voorouders nog een rol in kunnen spelen. Het erfelijk materiaal wordt niet door de koningin en/of de dar zelf geproduceerd of ontwikkeld of uitgebreid. Nee, zij kunnen niets anders dan doorgeven wat zij ook weer gekregen hebben.
De erffactoren, die bij de versmelting van zaadcel x eicel bij elkaar komen, blijven bij elkaar zolang de bij leeft. Voorafgaand aan de reductiedeling bij de nieuwe eicelvorming kan er echter door crossing-over verandering in de erffactoren komen. Door de grilligheid van de erffactoren ga ik er soms aan twijfelen of het wel mogelijk is, om met kennis van die erffactoren de eigenschappen van onze bijen te verbeteren of te vergroten. Toch blijft als een paal boven water staan, dat enige verwantschap de kans op gelukkige combinaties van erffactoren vergroot. Let wel, NIET GEGARANDEERD
Het schema probeert duidelijk te maken, waar achtereenvolgens die 50%, die 25%, die 12,5% en ook die 6,25% vandaan komen. In totaal zijn er in dit schema bij die 50% genen 8 + 8 = 16 voorouders betrokken.
Zo zien wij een steeds terugkerend beeld van ontstaan en vergaan, waarop weer een nieuw ontstaan volgt. Het verstrooit zich en komt weer tezamen. Maar nooit is het nieuwe een herhaling van het oude. De bij van gisteren is in de bij van vandaag gestorven, de bij van nu sterft in de bij van morgen. Het voorafgaande treedt in een veranderde samenstelling steeds weer te voorschijn.
Duidelijk is dat de invloed van ver verwijderde voorouders steeds minder wordt. Duidelijk is ook dat de grote cirkel in het midden nooit een bij kan voorstellen, waarvan de linker 50% precies gelijk is aan de rechter 50%. Alleen wanneer dit mogelijk zou zijn, kan er van een homozygote bij gesproken worden. De reductiedeling bij de vorming van een eicel drijft echter de spot met alle wiskundige berekeningen, die een superbij zouden moeten leveren, de gefantaseerde homozygoot. Jammer, maar die bestaat echt niet. Gelukkig maar!
Alles wat na de erfelijkheid nog een rol speelt zijn de milieuomstandigheden en dat is best nog heel wat. Denk maar eens aan de omstandigheden waaronder een nieuwe moer wordt opgetrokken b.v. uit zwermcellen, redcellen of teeltcellen of de situatie in pleeg- en bevruchtingsvolk(jes)en. Soms (vaak?) zijn deze milieuomstandigheden niet optimaal. Ook de huisvesting, drachtomstandigheden, verzorging, varroabehandeling en bedrijfsmethoden vallen hieronder.
!Piet